
The High Impact of Staphylococcus aureus Biofilm Culture Medium on In Vitro Outcomes of Antimicrobial Activity of Wound Antiseptics and Antibiotic

 , ,
, ,  , ,
, ,
Abstract
:
1. Introduction
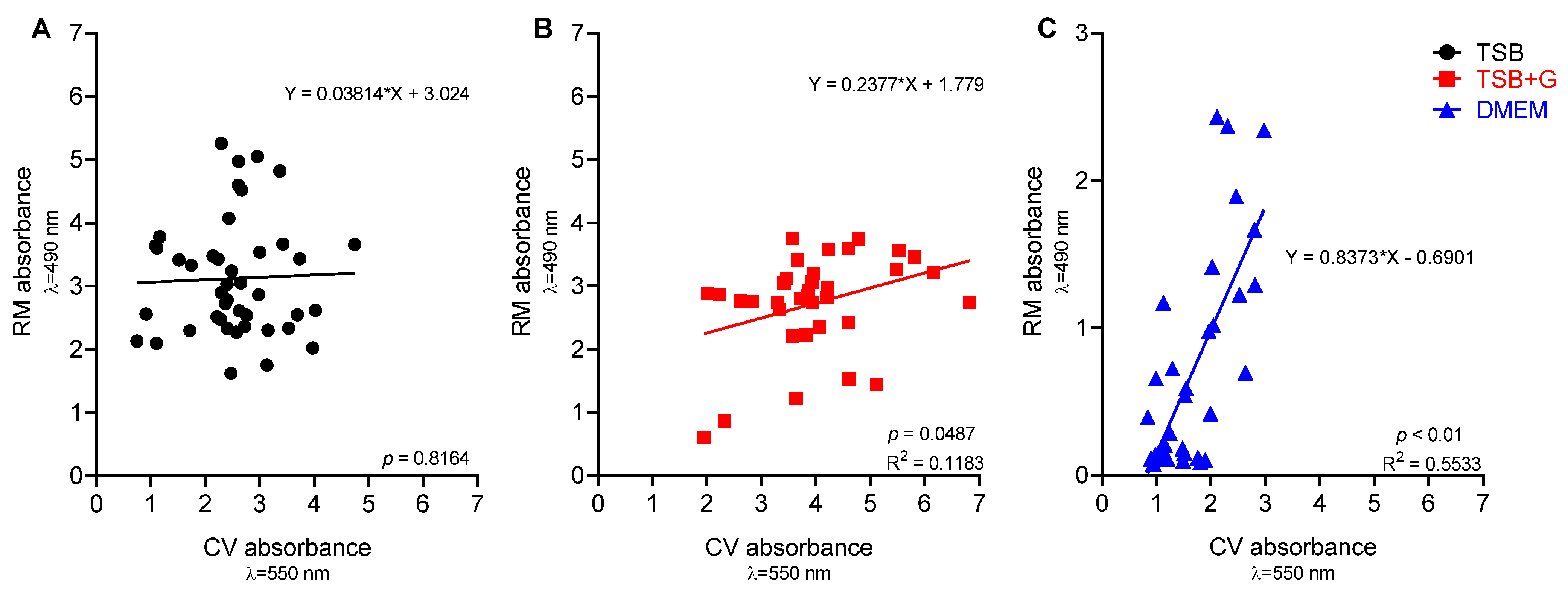
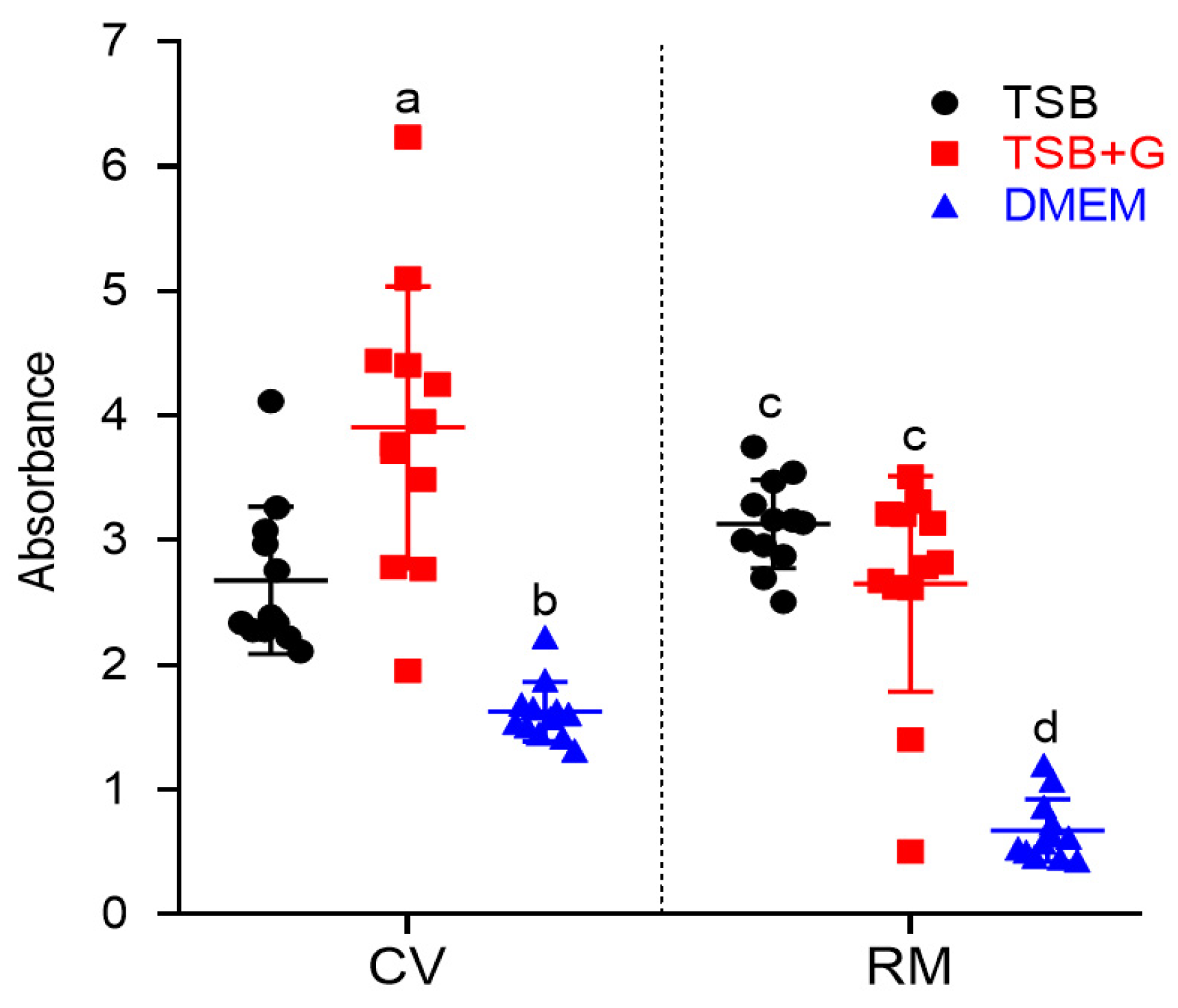
2. Results
3. Discussion
4. Materials and Methods
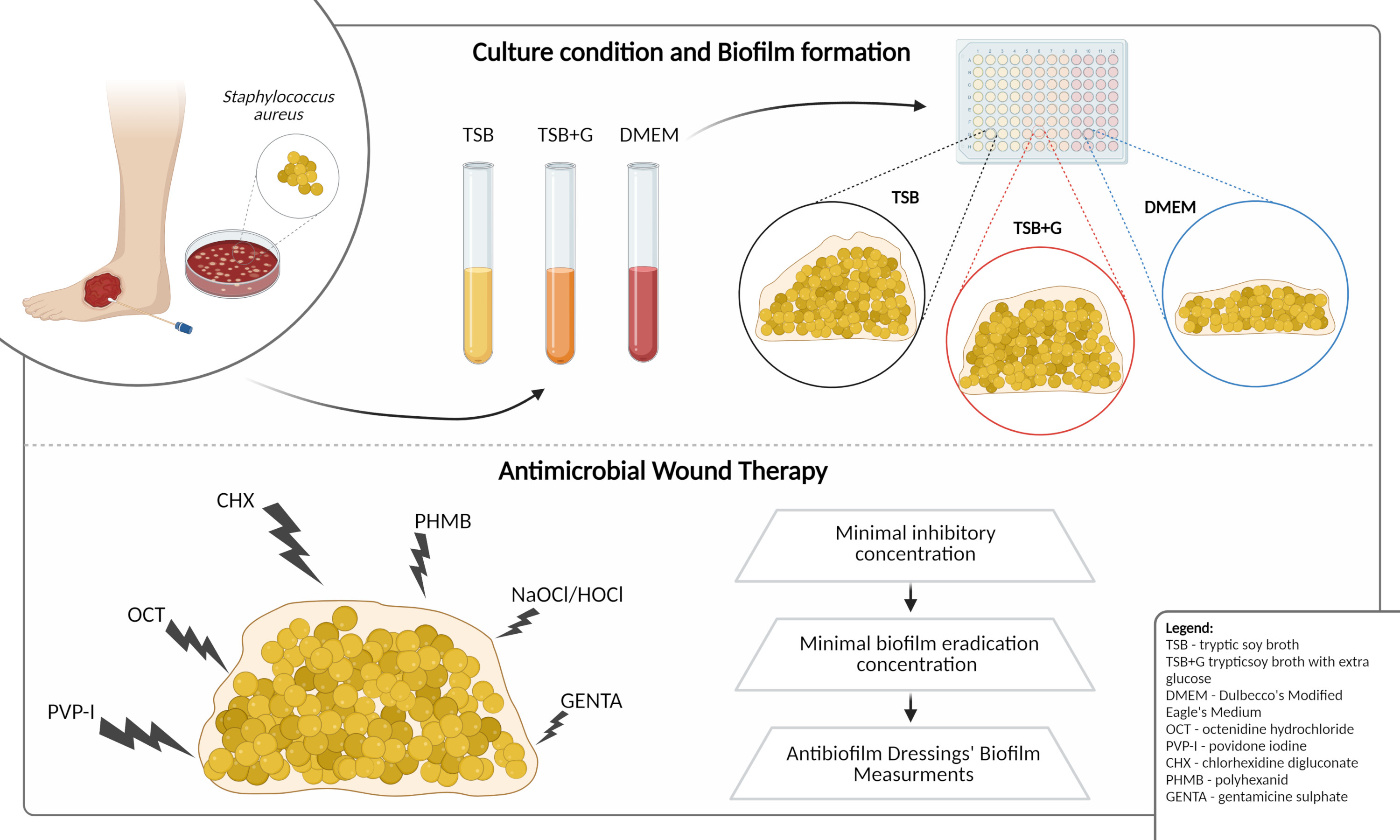
4.1. Microorganisms and Culture Conditions
- Tryptic soy broth (Biomaxima, Lublin, Poland), later referred to as TSB;
- Tryptic soy broth (Biomaxima, Lublin, Poland) supplemented with 1% glucose (w/v; Chempur, Piekary Slaskie, Poland), later referred to as TSB+G;
- Dulbecco’s modified Eagle’s medium high glucose (Biowest, Riverside, MO, USA; cat no. L0103) supplemented with 10% of fetal bovine serum (Biowest, Riverside, MO, USA), later referred to as DMEM.
4.2. Antimicrobials Applied
- Octenisept® (Schülke Mayr GmbH, Vienna, Austria), composed of 0.1% octenidine dihydrochloride, 2% phenoxyethanol, (3-amidpropyl cocoate) dimethylammonium acetate, sodium D gluconate, glycerol 85%, sodium chloride, sodium hydroxide, and purified water, later referred to as OCT;
- Braunol® (B. Braun, Melsungen, Hessen, Germany), composed 7.5% povidone-iodine with 10% available iodine, sodium dihydrogen phosphate dihydrate, sodium iodate, macrogol lauryl ether, sodium hydroxide, and purified water, later referred to as PVP-I;
- Prontosan® wound irrigation solution (B. Braun, Melsungen, Hessen, Germany), composed of purified water, 0.1% betaine surfactant, and 0.1% polyaminopropyl biguanide (polyhexanide), later referred to as PHMB;
- Chlorhexidine digluconate—pharmaceutical raw material (Fagron Pharma Cosmetics, Rotterdam, The Netherlands) to prepare a hydrous solution of final concentration 0.5%, later referred to as CHX;
- Granudacyn® Wound Irrigation Solution (Molnlycke Health Care AB, Göteborg, Sweden), composed of water, sodium chloride, 0.005% sodium hypochlorite, and 0.005% hypochlorous acid, later referred to as GRAN;
- Microdacyn60® Wound Care (Sonoma Pharmaceuticals, Inc, Petaluma, CA, USA), composed of super-oxidized water, sodium chloride, 0.004% sodium hypochlorite, and 0.004% hypochlorous acid, later referred to as MICR;
- Gentamicin sulfate powder (Pol-Aura, Dywity, Poland) to prepare a hydrous solution of final concentration of 0.2% later, later referred to as GENTA.
4.3. Assessment of Biofilm Biomass Level Performed Using Crystal Violet Dye (CV) in 96-Well Microtiter Plate
4.4. Assessment of Level of Biofilm Metabolic Activity Performed Using Richard’s Method (RM) in a 96-Well Microtiter Plate
4.5. Assessment of the Number of Colony-Forming Units Forming Biofilm Using Quantitative Culture and Richard’s Method
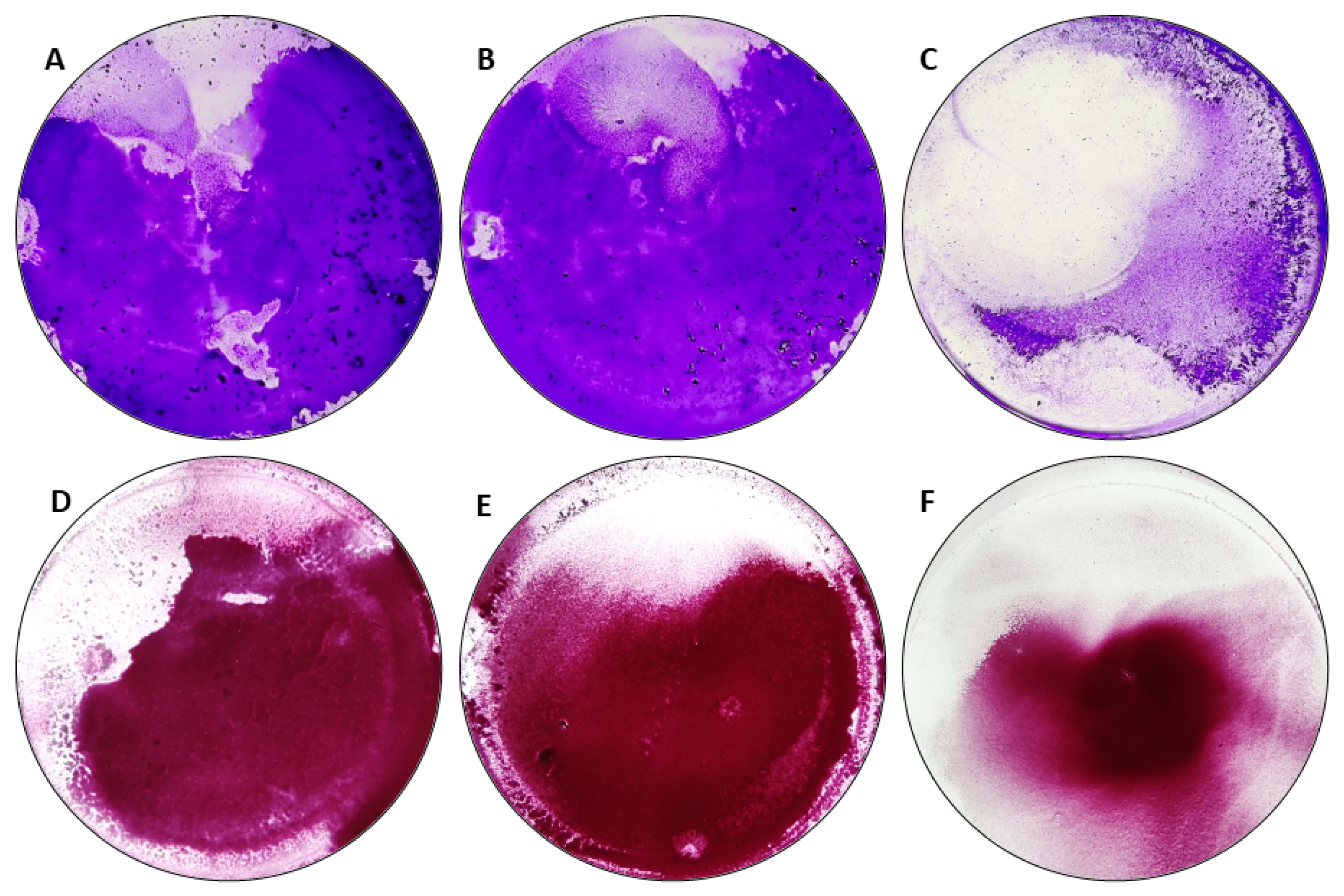
4.6. The Confirmation of Biofilm Formation by Means of Epifluorescent/Confocal Microscopy
4.7. Assessment of Antimicrobial Susceptibility of Analyzed Strains to Gentamicin Sulfate Using Disc Diffusion Method
4.8. Assessment of Minimal Inhibitory Concentration (MIC) of Applied Antimicrobials Using Spectrometric Assessment and RM in 96-Well Microtiter Plate
4.9. Assessment of Minimal Biofilm Eradication Concentration (MBEC) of Applied Antimicrobials Using Spectrometric Assessment and RM in a 96-Well Microtiter Plate
4.10. Modified Antibiofilm Dressing’s Activity Measurement (A.D.A.M.)
4.10.1. Preparation of Biocellulose Dressings
4.10.2. Antimicrobial Activity of Saturated Dressings
4.11. Statistical Analysis
5. Conclusions
- The application of various culture media changed the level of staphylococcal biofilm biomass and metabolic activity;
- The staphylococcal biofilms formed in DMEM displayed lower level of biomass and metabolic activity than biofilms formed in TSB and TSB+G;
- The effectiveness of inhibiting bacterial growth and biofilm formation of treatment agents was found to be dependent on type of grown biofilm and the medium applied;
- Results from in vitro studies should be scrutinized carefully with the stress put on the applied methodology.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Armstrong, D.G.; Bauer, K.; Bohn, G.; Carter, M.; Snyder, R.; Serena, T.E. Principles of Best Diagnostic Practice in Tissue Repair and Wound Healing: An Expert Consensus. Diagnostics 2020, 11, 50. [Google Scholar] [CrossRef] [PubMed]
- Wolcott, R.D.; Hanson, J.D.; Rees, E.J.; Koenig, L.D.; Phillips, C.D.; Wolcott, R.A.; Cox, S.B.; White, J.S. Analysis of the Chronic Wound Microbiota of 2,963 Patients by 16S RDNA Pyrosequencing. Wound Repair Regen. 2016, 24, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Shettigar, K.; Jain, S.; Bhat, D.V.; Acharya, R.; Ramachandra, L.; Satyamoorthy, K.; Murali, T.S. Virulence Determinants in Clinical Staphylococcus Aureus from Monomicrobial and Polymicrobial Infections of Diabetic Foot Ulcers. J. Med. Microbiol. 2016, 65, 1392–1404. [Google Scholar] [CrossRef] [PubMed]
- Moormeier, D.E.; Bayles, K.W. Staphylococcus aureus biofilm: A complex developmental organism. Mol. Microbiol. 2017, 104, 365–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omar, A.; Wright, J.B.; Schultz, G.; Burrell, R.; Nadworny, P. Microbial Biofilms and Chronic Wounds. Microorganisms 2017, 5, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, G.A.; Swogger, E.; Wolcott, R.; Pulcini, E.D.; Secor, P.; Sestrich, J.; Costerton, J.W.; Stewart, P.S. Biofilms in Chronic Wounds. Wound Repair Regen. Off. Publ. Wound Health Soc. Eur. Tissue Repair Soc. 2008, 16, 37–44. [Google Scholar] [CrossRef]
- Marano, R.J.; Wallace, H.J.; Wijeratne, D.; Fear, M.W.; Wong, H.S.; O’Handley, R. Secreted Biofilm Factors Adversely Affect Cellular Wound Healing Responses in Vitro. Sci. Rep. 2015, 5, 13296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosanquet, D.C.; Harding, K.G. Wound Duration and Healing Rates: Cause or Effect? Wound Repair Regen. 2014, 22, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.; Atkin, L.; Dissemond, J.; Hurlow, J.; Tan, Y.K.; Apelqvist, J.; James, G.; Salles, N.; Wu, J.; Tachi, M.; et al. Defying Hard-to-Heal Wounds with an Early Antibiofilm Intervention Strategy: ‘Wound Hygiene’. J. Wound Care 2019, 28, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Haesler, E.; Swanson, T.; Ousey, K.; Carville, K. Clinical Indicators of Wound Infection and Biofilm: Reaching International Consensus. J. Wound Care 2019, 28, s4–s12. [Google Scholar] [CrossRef] [PubMed]
- Kadam, S.; Madhusoodhanan, V.; Dhekane, R.; Bhide, D.; Ugale, R.; Tikhole, U.; Kaushik, K.S. Milieu Matters: An in Vitro Wound Milieu to Recapitulate Key Features of, and Probe New Insights into, Mixed-Species Bacterial Biofilms. Biofilm 2021, 3, 100047. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.; Dissemond, J.; Kim, S.; Willy, C.; Mayer, D.; Papke, R.; Tuchmann, F.; Assadian, O. Consensus on Wound Antisepsis: Update 2018. Skin Pharmacol. Physiol. 2018, 31, 28–58. [Google Scholar] [CrossRef] [PubMed]
- Broxton, P.; Woodcock, P.M.; Heatley, F.; Gilbert, P. Interaction of Some Polyhexamethylene Biguanides and Membrane Phospholipids in Escherichia Coli. J. Appl. Bacteriol. 1984, 57, 115–124. [Google Scholar] [CrossRef]
- Chindera, K.; Mahato, M.; Sharma, A.K.; Horsley, H.; Kloc-Muniak, K.; Kamaruzzaman, N.F.; Kumar, S.; McFarlane, A.; Stach, J.; Bentin, T.; et al. The Antimicrobial Polymer PHMB Enters Cells and Selectively Condenses Bacterial Chromosomes. Sci. Rep. 2016, 6, 23121. [Google Scholar] [CrossRef] [PubMed]
- Brill, F.H.H.; Gabriel, H. Is Polyhexamethylene-Guanidine Hydrochloride (PHMGH) Sporicidal? A Critical Review. J. Med. Microbiol. 2015, 64, 307–308. [Google Scholar] [CrossRef] [Green Version]
- Machuca, J.; Lopez-Rojas, R.; Fernandez-Cuenca, F.; Pascual, Á. Comparative Activity of a Polyhexanide-Betaine Solution against Biofilms Produced by Multidrug-Resistant Bacteria Belonging to High-Risk Clones. J. Hosp. Infect. 2019, 103, e92–e96. [Google Scholar] [CrossRef] [PubMed]
- Hübner, N.-O.; Siebert, J.; Kramer, A. Octenidine Dihydrochloride, a Modern Antiseptic for Skin, Mucous Membranes and Wounds. Skin Pharmacol. Physiol. 2010, 23, 244–258. [Google Scholar] [CrossRef] [PubMed]
- Krasowski, G.; Junka, A.; Paleczny, J.; Czajkowska, J.; Makomaska-Szaroszyk, E.; Chodaczek, G.; Majkowski, M.; Migdał, P.; Fijałkowski, K.; Kowalska-Krochmal, B.; et al. In Vitro Evaluation of Polihexanide, Octenidine and NaClO/HClO-Based Antiseptics against Biofilm Formed by Wound Pathogens. Membranes 2021, 11, 62. [Google Scholar] [CrossRef] [PubMed]
- Hugo, W.B.; Longworth, A.R. The Effect of Chlorhexidine on the Electrophoretic Mobility, Cytoplasmic Constituents, Dehydrogenase Activity and Cell Walls of Escherichia Coli and Staphylococcus Aureus. J. Pharm. Pharmacol. 1966, 18, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Htun, H.L.; Hon, P.Y.; Holden, M.T.G.; Ang, B.; Chow, A. Chlorhexidine and Octenidine Use, Carriage of Qac Genes, and Reduced Antiseptic Susceptibility in Methicillin-Resistant Staphylococcus Aureus Isolates from a Healthcare Network. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2019, 25, 1154.e1–1154.e7. [Google Scholar] [CrossRef] [Green Version]
- Bigliardi, P.L.; Alsagoff, S.A.L.; El-Kafrawi, H.Y.; Pyon, J.-K.; Wa, C.T.C.; Villa, M.A. Povidone Iodine in Wound Healing: A Review of Current Concepts and Practices. Int. J. Surg. 2017, 44, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Hill, K.E.; Malic, S.; McKee, R.; Rennison, T.; Harding, K.G.; Williams, D.W.; Thomas, D.W. An in Vitro Model of Chronic Wound Biofilms to Test Wound Dressings and Assess Antimicrobial Susceptibilities. J. Antimicrob. Chemother. 2010, 65, 1195–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oduwole, K.O.; Glynn, A.A.; Molony, D.C.; Murray, D.; Rowe, S.; Holland, L.M.; McCormack, D.J.; O’Gara, J.P. Anti-Biofilm Activity of Sub-Inhibitory Povidone-Iodine Concentrations against Staphylococcus Epidermidis and Staphylococcus Aureus. J. Orthop. Res. 2010, 28, 1252–1256. [Google Scholar] [CrossRef] [PubMed]
- Anagnostopoulos, A.G.; Rong, A.; Miller, D.; Tran, A.Q.; Head, T.; Lee, M.C.; Lee, W.W. 0.01% Hypochlorous Acid as an Alternative Skin Antiseptic: An In Vitro Comparison. Dermatol. Surg. Off. Publ. Am. Soc. Dermatol. Surg. Al 2018, 44, 1489–1493. [Google Scholar] [CrossRef]
- Herruzo, R.; Herruzo, I. Antimicrobial Efficacy of a Very Stable Hypochlorous Acid Formula Compared with Other Antiseptics Used in Treating Wounds: In-Vitro Study on Micro-Organisms with or without Biofilm. J. Hosp. Infect. 2020, 105, 289–294. [Google Scholar] [CrossRef]
- Rembe, J.-D.; Huelsboemer, L.; Plattfaut, I.; Besser, M.; Stuermer, E.K. Antimicrobial Hypochlorous Wound Irrigation Solutions Demonstrate Lower Anti-Biofilm Efficacy Against Bacterial Biofilm in a Complex in-Vitro Human Plasma Biofilm Model (HpBIOM) Than Common Wound Antimicrobials. Front. Microbiol. 2020, 11, 564513. [Google Scholar] [CrossRef] [PubMed]
- Severing, A.-L.; Rembe, J.-D.; Koester, V.; Stuermer, E.K. Safety and Efficacy Profiles of Different Commercial Sodium Hypochlorite/Hypochlorous Acid Solutions (NaClO/HClO): Antimicrobial Efficacy, Cytotoxic Impact and Physicochemical Parameters in Vitro. J. Antimicrob. Chemother. 2019, 74, 365–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, Y.; Wu, P.; Chen, B. Update on New Medicinal Applications of Gentamicin: Evidence-Based Review. J. Formos. Med. Assoc. 2014, 113, 72–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maczynska, B.; Secewicz, A.; Smutnicka, D.; Szymczyk, P.; Dudek-Wicher, R.; Junka, A.; Bartoszewicz, M. In Vitro Efficacy of Gentamicin Released from Collagen Sponge in Eradication of Bacterial Biofilm Preformed on Hydroxyapatite Surface. PLoS ONE 2019, 14, e0217769. [Google Scholar] [CrossRef] [PubMed]
- Cutting, K.F. Wound Exudate: Composition and Functions. Br. J. Community Nurs. 2003, 8, S4–S9. [Google Scholar] [CrossRef] [PubMed]
- Power, G.; Moore, Z.; O’Connor, T. Measurement of PH, Exudate Composition and Temperature in Wound Healing: A Systematic Review. J. Wound Care 2017, 26, 381–397. [Google Scholar] [CrossRef]
- Trengove, N.J.; Langton, S.R.; Stacey, M.C. Biochemical Analysis of Wound Fluid from Nonhealing and Healing Chronic Leg Ulcers. Wound Repair Regen. Off. Publ. Wound Health Soc. Eur. Tissue Repair Soc. 1996, 4, 234–239. [Google Scholar] [CrossRef]
- Trivedi, U.; Madsen, J.S.; Rumbaugh, K.P.; Wolcott, R.D.; Burmølle, M.; Sørensen, S.J. A Post-Planktonic Era of in Vitro Infectious Models: Issues and Changes Addressed by a Clinically Relevant Wound like Media. Crit. Rev. Microbiol. 2017, 43, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, G.; Dilhari, A.; Gayani, B.; Kottegoda, N.; Samaranayake, L.; Weerasekera, M. Influence of Laboratory Culture Media on in Vitro Growth, Adhesion, and Biofilm Formation of Pseudomonas Aeruginosa and Staphylococcus Aureus. Med. Princ. Pract. Int. J. Kuwait Univ. Health Sci. Cent. 2019, 28, 28–35. [Google Scholar] [CrossRef]
- Junka, A.F.; Żywicka, A.; Szymczyk, P.; Dziadas, M.; Bartoszewicz, M.; Fijałkowski, K.A.D.A.M. Test (Antibiofilm Dressing’s Activity Measurement)—Simple Method for Evaluating Anti-Biofilm Activity of Drug-Saturated Dressings against Wound Pathogens. J. Microbiol. Methods 2017, 143, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Werthén, M.; Henriksson, L.; Jensen, P.Ø.; Sternberg, C.; Givskov, M.; Bjarnsholt, T. An in Vitro Model of Bacterial Infections in Wounds and Other Soft Tissues. APMIS Acta Pathol. Microbiol. Immunol. Scand. 2010, 118, 156–164. [Google Scholar] [CrossRef]
- Pouget, C.; Gustave, C.-A.; Ngba-Essebe, C.; Laurent, F.; Lemichez, E.; Tristan, A.; Sotto, A.; Dunyach-Rémy, C.; Lavigne, J.-P. Adaptation of Staphylococcus Aureus in a Medium Mimicking a Diabetic Foot Environment. Toxins 2021, 13, 230. [Google Scholar] [CrossRef] [PubMed]
- Pitten, F.-A.; Werner, H.-P.; Kramer, A. A Standardized Test to Assess the Impact of Different Organic Challenges on the Antimicrobial Activity of Antiseptics. J. Hosp. Infect. 2003, 55, 108–115. [Google Scholar] [CrossRef]
- Junker, J.P.E.; Caterson, E.J.; Eriksson, E. The Microenvironment of Wound Healing. J. Craniofac. Surg. 2013, 24, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Junka, A.; Wojtowicz, W.; Ząbek, A.; Krasowski, G.; Smutnicka, D.; Bakalorz, B.; Boruta, A.; Dziadas, M.; Młynarz, P.; Sedghizadeh, P.P.; et al. Metabolic Profiles of Exudates from Chronic Leg Ulcerations. J. Pharm. Biomed. Anal. 2017, 137, 13–22. [Google Scholar] [CrossRef]
- Alonso, B.; Lucio, J.; Pérez-Granda, M.J.; Cruces, R.; Sánchez-Carrillo, C.; Bouza, E.; Guembe, M. Does Biomass Production Correlate with Metabolic Activity in Staphylococcus Aureus? J. Microbiol. Methods 2016, 131, 110–112. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Liang, Y.; Lin, S.; Chen, D.; Li, B.; Li, L.; Deng, Y. Crystal Violet and XTT Assays on Staphylococcus Aureus Biofilm Quantification. Curr. Microbiol. 2016, 73, 474–482. [Google Scholar] [CrossRef]
- Sousa, C.; Teixeira, P.; Oliveira, R. The Role of Extracellular Polymers on Staphylococcus Epidermidis Biofilm Biomass and Metabolic Activity. J. Basic Microbiol. 2009, 49, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Waldrop, R.; McLaren, A.; Bse, F.C.; McLemore, R. Biofilm Growth Has a Threshold Response to Glucose in Vitro. Clin. Orthop. 2014, 472, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lade, H.; Park, J.H.; Chung, S.H.; Kim, I.H.; Kim, J.-M.; Joo, H.-S.; Kim, J.-S. Biofilm Formation by Staphylococcus Aureus Clinical Isolates Is Differentially Affected by Glucose and Sodium Chloride Supplemented Culture Media. J. Clin. Med. 2019, 8, 1853. [Google Scholar] [CrossRef] [Green Version]
- Tan, L.; Li, S.R.; Jiang, B.; Hu, X.M.; Li, S. Therapeutic Targeting of the Staphylococcus Aureus Accessory Gene Regulator (Agr) System. Front. Microbiol. 2018, 9, 55. [Google Scholar] [CrossRef]
- Vuong, C.; Saenz, H.L.; Götz, F.; Otto, M. Impact of the Agr Quorum-Sensing System on Adherence to Polystyrene in Staphylococcus Aureus. J. Infect. Dis. 2000, 182, 1688–1693. [Google Scholar] [CrossRef] [Green Version]
- Boles, B.R.; Horswill, A.R. Agr-Mediated Dispersal of Staphylococcus Aureus Biofilms. PLoS Pathog. 2008, 4, e1000052. [Google Scholar] [CrossRef]
- Croes, S.; Deurenberg, R.H.; Boumans, M.-L.L.; Beisser, P.S.; Neef, C.; Stobberingh, E.E. Staphylococcus Aureus Biofilm Formation at the Physiologic Glucose Concentration Depends on the S. Aureus Lineage. BMC Microbiol. 2009, 9, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, Y.; Xue, T.; Cao, L.; Zhao, L.; Sun, H.; Sun, B. Staphylococcus Aureus Glucose-Induced Biofilm Accessory Proteins, GbaAB, Influence Biofilm Formation in a PIA-Dependent Manner. Int. J. Med. Microbiol. IJMM 2014, 304, 603–612. [Google Scholar] [CrossRef]
- O’Gara, J.P. Ica and beyond: Biofilm Mechanisms and Regulation in Staphylococcus Epidermidis and Staphylococcus Aureus. FEMS Microbiol. Lett. 2007, 270, 179–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, E.; Pozzi, C.; Houston, P.; Smyth, D.; Humphreys, H.; Robinson, D.A.; O’Gara, J.P. Association between Methicillin Susceptibility and Biofilm Regulation in Staphylococcus Aureus Isolates from Device-Related Infections. J. Clin. Microbiol. 2007, 45, 1379–1388. [Google Scholar] [CrossRef] [Green Version]
- Ghasemian, A.; Najar Peerayeh, S.; Bakhshi, B.; Mirzaee, M. Comparison of Biofilm Formation between Methicillin-Resistant and Methicillin-Susceptible Isolates of Staphylococcus Aureus. Iran. Biomed. J. 2016, 20, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Senobar Tahaei, S.A.; Stájer, A.; Barrak, I.; Ostorházi, E.; Szabó, D.; Gajdács, M. Correlation Between Biofilm-Formation and the Antibiotic Resistant Phenotype in Staphylococcus Aureus Isolates: A Laboratory-Based Study in Hungary and a Review of the Literature. Infect. Drug Resist. 2021, 14, 1155–1168. [Google Scholar] [CrossRef] [PubMed]
- Halsey, C.R.; Lei, S.; Wax, J.K.; Lehman, M.K.; Nuxoll, A.S.; Steinke, L.; Sadykov, M.; Powers, R.; Fey, P.D. Amino Acid Catabolism in Staphylococcus Aureus and the Function of Carbon Catabolite Repression. mBio 2017, 8, e01434-16. [Google Scholar] [CrossRef] [Green Version]
- Bodini, S.; Nunziangeli, L.; Santori, F. Influence of Amino Acids on Low-Density Escherichia Coli Responses to Nutrient Downshifts. J. Bacteriol. 2007, 189, 3099–3105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolodkin-Gal, I.; Romero, D.; Cao, S.; Clardy, J.; Kolter, R.; Losick, R. D-Amino Acids Trigger Biofilm Disassembly. Science 2010, 328, 627–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochbaum, A.I.; Kolodkin-Gal, I.; Foulston, L.; Kolter, R.; Aizenberg, J.; Losick, R. Inhibitory Effects of D-Amino Acids on Staphylococcus Aureus Biofilm Development. J. Bacteriol. 2011, 193, 5616–5622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zara, G.; Bou Zeidan, M.; Fancello, F.; Sanna, M.L.; Mannazzu, I.; Budroni, M.; Zara, S. The Administration of L-Cysteine and l-Arginine Inhibits Biofilm Formation in Wild-Type Biofilm-Forming Yeast by Modulating FLO11 Gene Expression. Appl. Microbiol. Biotechnol. 2019, 103, 7675–7685. [Google Scholar] [CrossRef]
- Dobay, O.; Laub, K.; Stercz, B.; Kéri, A.; Balázs, B.; Tóthpál, A.; Kardos, S.; Jaikumpun, P.; Ruksakiet, K.; Quinton, P.M.; et al. Bicarbonate Inhibits Bacterial Growth and Biofilm Formation of Prevalent Cystic Fibrosis Pathogens. Front. Microbiol. 2018, 9, 2245. [Google Scholar] [CrossRef] [Green Version]
- Jaikumpun, P.; Ruksakiet, K.; Stercz, B.; Pállinger, É.; Steward, M.; Lohinai, Z.; Dobay, O.; Zsembery, Á. Antibacterial Effects of Bicarbonate in Media Modified to Mimic Cystic Fibrosis Sputum. Int. J. Mol. Sci. 2020, 21, 8614. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.K.; Rao, T.S. Effect of Calcium on Staphylococcus Aureus Biofilm Architecture: A Confocal Laser Scanning Microscopic Study. Colloids Surf. B Biointerfaces 2013, 103, 448–454. [Google Scholar] [CrossRef]
- Abraham, N.M.; Lamlertthon, S.; Fowler, V.G.; Jefferson, K.K. Chelating Agents Exert Distinct Effects on Biofilm Formation in Staphylococcus Aureus Depending on Strain Background: Role for Clumping Factor B. J. Med. Microbiol. 2012, 61, 1062–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrizubieta, M.J.; Toledo-Arana, A.; Amorena, B.; Penadés, J.R.; Lasa, I. Calcium Inhibits Bap-Dependent Multicellular Behavior in Staphylococcus Aureus. J. Bacteriol. 2004, 186, 7490–7498. [Google Scholar] [CrossRef] [Green Version]
- Dydak, K.; Junka, A.; Dydak, A.; Brożyna, M.; Paleczny, J.; Fijalkowski, K.; Kubielas, G.; Aniołek, O.; Bartoszewicz, M. In Vitro Efficacy of Bacterial Cellulose Dressings Chemisorbed with Antiseptics against Biofilm Formed by Pathogens Isolated from Chronic Wounds. Int. J. Mol. Sci. 2021, 22, 3996. [Google Scholar] [CrossRef]
- FDA. Drug Safety Communication FDA Warns about Rare but Serious Allergic Reactions with the Skin Antiseptic Chlorhexidine Gluconate. 2019. Available online: https://www.fda.gov/media/102986/download (accessed on 1 January 2021).
- Schmidt, J.; Zyba, V.; Jung, K.; Rinke, S.; Haak, R.; Mausberg, R.F.; Ziebolz, D. Cytotoxic Effects of Octenidine Mouth Rinse on Human Fibroblasts and Epithelial Cells—An in Vitro Study. Drug Chem. Toxicol. 2016, 39, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Faria, G.; Celes, M.R.N.; De Rossi, A.; Silva, L.A.B.; Silva, J.S.; Rossi, M.A. Evaluation of Chlorhexidine Toxicity Injected in the Paw of Mice and Added to Cultured L929 Fibroblasts. J. Endod. 2007, 33, 715–722. [Google Scholar] [CrossRef]
- Kapalschinski, N.; Seipp, H.M.; Kückelhaus, M.; Harati, K.K.; Kolbenschlag, J.J.; Daigeler, A.; Jacobsen, F.; Lehnhardt, M.; Hirsch, T. Albumin Reduces the Antibacterial Efficacy of Wound Antiseptics against Staphylococcus Aureus. J. Wound Care 2017, 26, 184–187. [Google Scholar] [CrossRef]
- Radischat, N.; Augustin, M.; Herberger, K.; Wille, A.; Goroncy-Bermes, P. Influence of Human Wound Exudate on the Bactericidal Efficacy of Antiseptic Agents in Quantitative Suspension Tests on the Basis of European Standards (DIN EN 13727). Int. Wound J. 2020, 17, 781–789. [Google Scholar] [CrossRef]
- Junka, A.; Bartoszewicz, M.; Dziadas, M.; Szymczyk, P.; Dydak, K.; Żywicka, A.; Owczarek, A.; Bil-Lula, I.; Czajkowska, J.; Fijałkowski, K. Application of Bacterial Cellulose Experimental Dressings Saturated with Gentamycin for Management of Bone Biofilm in Vitro and Ex Vivo. J. Biomed. Mater. Res. B Appl. Biomater. 2020, 108, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, G.; Dey, D.; Das, S.; Banerjee, A. Exposure to Sub-Inhibitory Concentrations of Gentamicin, Ciprofloxacin and Cefotaxime Induces Multidrug Resistance and Reactive Oxygen Species Generation in Meticillin-Sensitive Staphylococcus Aureus. J. Med. Microbiol. 2017, 66, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Cattelaens, J.; Turco, L.; Berclaz, L.M.; Huelsse, B.; Hitzl, W.; Vollkommer, T.; Bodenschatz, K.J. The Impact of a Nanocellulose-Based Wound Dressing in the Management of Thermal Injuries in Children: Results of a Retrospective Evaluation. Life Basel Switz. 2020, 10, 212. [Google Scholar] [CrossRef] [PubMed]
- Krzyżek, P.; Gościniak, G.; Fijałkowski, K.; Migdał, P.; Dziadas, M.; Owczarek, A.; Czajkowska, J.; Aniołek, O.; Junka, A. Potential of Bacterial Cellulose Chemisorbed with Anti-Metabolites, 3-Bromopyruvate or Sertraline, to Fight against Helicobacter Pylori Lawn Biofilm. Int. J. Mol. Sci. 2020, 21, 9507. [Google Scholar] [CrossRef]
- Krasowski, G.; Wicher-Dudek, R.; Paleczny, J.; Bil-Lula, I.; Fijałkowski, K.; Sedghizadeh, P.P.; Szymczyk, P.; Dudek, B.; Bartoszewicz, M.; Junka, A. Potential of Novel Bacterial Cellulose Dressings Chemisorbed with Antiseptics for the Treatment of Oral Biofilm Infections. Appl. Sci. 2019, 9, 5321. [Google Scholar] [CrossRef] [Green Version]
- Latka, A.; Drulis-Kawa, Z. Advantages and Limitations of Microtiter Biofilm Assays in the Model of Antibiofilm Activity of Klebsiella Phage KP34 and Its Depolymerase. Sci. Rep. 2020, 10, 20338. [Google Scholar] [CrossRef]
- Thaarup, I.C.; Bjarnsholt, T. Current In Vitro Biofilm-Infected Chronic Wound Models for Developing New Treatment Possibilities. Adv. Wound Care 2021, 10, 91–102. [Google Scholar] [CrossRef] [PubMed]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 11.0. 2021. Available online: https://eucast.org/clinical_breakpoints/ (accessed on 1 January 2021).
- Ciechanska, D.; Wietecha, J.; Kaźmierczak, D.; Kazimierczak, J. Biosynthesis of Modified Bacterial Cellulose in a Tubular Form. Fibres Text. East. Eur. 2010, 82, 98–104. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance | Comparison of MIC Values in Specific Media | ||||||
|---|---|---|---|---|---|---|---|
| TSB vs. TSB+G | TSB vs. DMEM | TSB+G vs. DMEM | |||||
| OCT | ns | ↑ | ↓ | *** | ↑ | ↓ | *** |
| PVP-I | ns | ↓ | ↑ | ** | ↓ | ↑ | *** |
| CHX | ns | ↑ | ↓ | * | ↑ | ↓ | * |
| PHMB | ns | ↑ | ↓ | *** | ↑ | ↓ | *** |
| GENTA | ns | ns | ns | ||||
| Substance | Comparison of MBEC Values in Specific Media | ||||||
|---|---|---|---|---|---|---|---|
| TSB vs. TSB+G | TSB vs. DMEM | TSB+G vs. DMEM | |||||
| OCT | ns | ↑ | ↓ | *** | ↑ | ↓ | ** |
| PVP-I | ns | ↑ | ↓ | *** | ↑ | ↓ | * |
| CHX | ns | ↑ | ↓ | ** | ↑ | ↓ | ** |
| PHMB | ns | ↑ | ↓ | *** | ↑ | ↓ | ** |
| (A) Tryptic Soy Broth (TSB) | g/L |
| Proteins | |
| Pancreatic digest of casein | 17 |
| Peptic digest of soybean | 3 |
| Inorganic salts | |
| Dipotassium hydrogen phosphate | 2.5 |
| Sodium chloride | 5 |
| Other components | |
| Glucose monohydrate | 2.5/10 * |
| (B) Dulbecco’s Modified Eagle’s Medium (DMEM) High Glucose L0103-500 Liquid | g/L |
| Amino acids | |
| Glycine | 0.03 |
| L-Alanyl-L-glutamine (glutamine stable) | 0.862 |
| L-Arginine monohydrochloride | 0.084 |
| L-Cystine dihydrochloride | 0.0626 |
| L-Histidine monohydrochloride monohydrate | 0.042 |
| L-Isoleucine | 0.105 |
| L-Leucine | 0.105 |
| L-Lysine monohydrochloride | 0.146 |
| L-Methionine | 0.03 |
| L-Phenylalanine | 0.066 |
| L-Serine | 0.042 |
| L-Threonine | 0.095 |
| L-Tryptophan | 0.016 |
| L-Tyrosine disodium salt dihydrate | 0.10379 |
| L-Valine | 0.094 |
| Inorganic salts | |
| Calcium chloride dihydrate | 0.265 |
| Ferric nitrate nonahydrate | 0.0001 |
| Magnesium sulfate anhydrous | 0.09767 |
| Potassium chloride | 0.4 |
| Sodium bicarbonate | 3.7 |
| Sodium chloride | 6.4 |
| Sodium phosphate monobasic anhydrous | 0.109 |
| Vitamins | |
| Choline chloride | 0.004 |
| D-Ca pantothenate | 0.004 |
| Folic acid | 0.004 |
| Myo-inositol | 0.0072 |
| Nicotinamide | 0.004 |
| Pyridoxal hydrochloride | 0.004 |
| Riboflavine | 0.0004 |
| Thiamine hydrochloride | 0.004 |
| Other components | |
| D-Glucose anhydrous | 4.5 |
| Phenol red solution salt | 0.0159 |
| Sodium pyruvate | 0.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paleczny, J.; Junka, A.; Brożyna, M.; Dydak, K.; Oleksy-Wawrzyniak, M.; Ciecholewska-Juśko, D.; Dziedzic, E.; Bartoszewicz, M. The High Impact of Staphylococcus aureus Biofilm Culture Medium on In Vitro Outcomes of Antimicrobial Activity of Wound Antiseptics and Antibiotic. Pathogens 2021, 10, 1385. https://doi.org/10.3390/pathogens10111385
Paleczny J, Junka A, Brożyna M, Dydak K, Oleksy-Wawrzyniak M, Ciecholewska-Juśko D, Dziedzic E, Bartoszewicz M. The High Impact of Staphylococcus aureus Biofilm Culture Medium on In Vitro Outcomes of Antimicrobial Activity of Wound Antiseptics and Antibiotic. Pathogens. 2021; 10(11):1385. https://doi.org/10.3390/pathogens10111385
Chicago/Turabian StylePaleczny, Justyna, Adam Junka, Malwina Brożyna, Karolina Dydak, Monika Oleksy-Wawrzyniak, Daria Ciecholewska-Juśko, Ewelina Dziedzic, and Marzenna Bartoszewicz. 2021. "The High Impact of Staphylococcus aureus Biofilm Culture Medium on In Vitro Outcomes of Antimicrobial Activity of Wound Antiseptics and Antibiotic" Pathogens 10, no. 11: 1385. https://doi.org/10.3390/pathogens10111385